视神经损伤
-
Figure 1|Major changes observed in microglial cells at different time points after OHT induction in the unilateral laser-induced OHT mouse model.
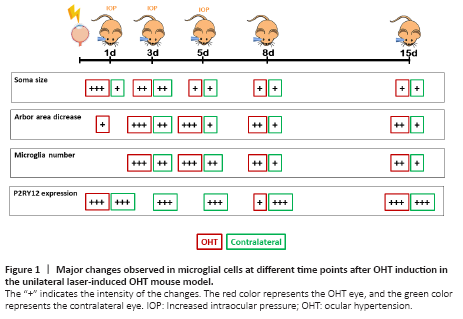
Considering the evidence of the involvement of the immune system in glaucomatous optic neuropathy and the important role of glial cell activation in RGC survival or death, an exhaustive study of how microglial activation progresses over time after OHT induction, in the laser-induced OHT model, would be a logical and interesting step. A study has examined the evolution of microglial activation after unilateral laser-induced OHT at different time points (1, 3, 5, 8, and 15 days) after OHT induction (Ramírez et al., 2020). Two markers have been used to identify microglia: Iba-1 and P2RY12. Iba-1 allows for the analysis of the morphological features of microglia, and the purinergic P2RY12 receptor allows for the differentiation of resident microglia (P2RY12+) from monocytes, infiltrating macrophages, or dendritic cells, which do not express this marker. In this study, an analysis is conducted on the different microglial morphological changes, such as changes of the soma size, the arbor area, the number of vertical processes, and the number of microglial cells, in all retinal layers in which microglia are present (the outer segment photoreceptor layer, and plexiform layers (outer and inner), and nerve fiber layer/ganglion cell layer (NFL-GCL)). Additionally, the P2RY12 expression of the microglia is analyzed (Figure 1). At 24 hours after unilateral laser OHT induction, the microglia showed signs of activation in all retinal layers where they were located (de Hoz et al., 2018; Ramírez et al., 2020). These signs included an increase in the soma size, shortening and reorientation of the processes, and the presence of macrophage amoebic cells, both in the OHT eye and in the normotensive contralateral eye, although it was more intense in the OHT eye. However, at this time, there was no increase in the number of microglial cells, although there was an increase in the area occupied by the microglia in the NFL-GCL layer, which could be due to migratory phenomena from other layers and an increase in the size of these cells, rather than an increase in their number (de Hoz et al., 2018). The activation of the microglial cells may be related to the increase in the IOP, which remains high in this model from day 1 to day 5, until it drops to normal values at 7 days. This IOP increase could produce damage in the RGCs, which would induce the activation of the microglial cells (Gallego et al., 2012; de Hoz et al., 2013, 2018; Rojas et al., 2014; Ramírez et al., 2020). The activated microglia would produce pro-inflammatory factors and chemokines that would induce the disruption of the BRB (Ramírez et al., 2015). At 24 hours after OHT induction, the microglia did not show an upregulation of MHC-II (de Hoz et al., 2018). However, there were MHC-II+ rounded cells, which were not microglia, because they did not express P2RY12, and could be monocytes infiltrated from the bloodstream due to the disruption of the BRB (Ramírez et al., 2020). These MHCII+ cells could induce the transformation of ramified microglial cells into amoeboid microglia, which would phagocytize the cells that had entered from the bloodstream (de Hoz et al., 2018). In addition, the microglia in both the OHT and contralateral eyes (more frequently in the OHT eyes) change their arrangement from parallel to the retinal surface to perpendicular, sending processes towards the neighboring microglial plexuses and connecting them. This could help in the transmission of information from the layers where the alteration of the BRB is taking place to the other retinal layers, thus contributing to microglial activation (de Hoz et al., 2018).