视神经损伤
-
Figure 2|Expression of sox11b in intact and injured zebrafish retinas.
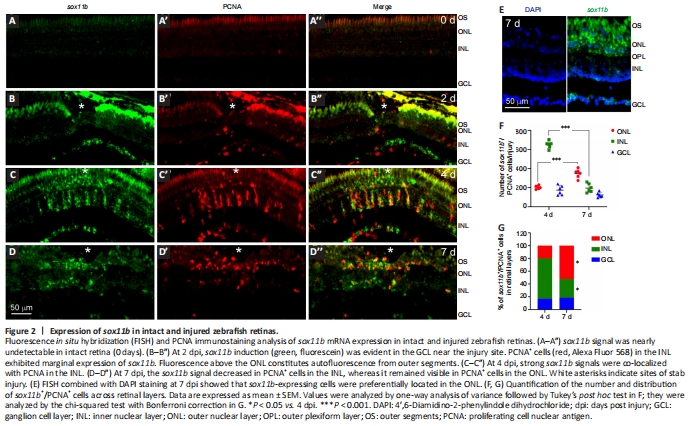
We have previously shown that after stab injury, sox11b is induced in the injured zebrafish retina and co-localizes with inner nuclear layer (INL) BrdU+ cells at 4 dpi (Mu et al., 2017). To better understand the expression pattern of sox11b, FISH using a sox11b antisense probe followed by PCNA immunofluorescence was performed in intact and injured zebrafish retinas at 2, 4 and 7 dpi. FISH showed that the sox11b mRNA signal was almost undetectable in intact retinas (Figure 2A–A''). At 2 dpi, sox11b was induced in the ganglion cell layer (GCL) but was nearly undetectable in the INL (Figure 2B–B''). Considering that MG entered the cell cycle at 2 days after stab injury (Fausett and Goldman, 2006; Zhang et al., 2020), our findings suggest that sox11b was induced in retinal ganglion cells (RGCs) rather than proliferating MG at 2 dpi. At 4 dpi, robust sox11b signals that co-localized with PCNA+ cell clusters were observed in the INL (Figure 2C–C''). Because MGPCs reportedly constitute almost all proliferating cells in the INL at 4 dpi (Fausett and Goldman, 2006), we inferred that sox11b was predominantly expressed in the progenitor cells. At 7 dpi, there were fewer sox11b+/PCNA+ cells in the INL (compared with 4 dpi); instead, sox11b+/PCNA+ cells were preferentially located in the outer nuclear layer (ONL) (Figure 2D–D'', and E–G). Thus, many of the sox11b-expressing MGPCs presumably migrated to the ONL at later time points, where they contribute to photoreceptor regeneration.
Figure 3|Sox11b knockdown does not affect MGPC cell number but causes changes in nuclear morphology at 4 dpi.
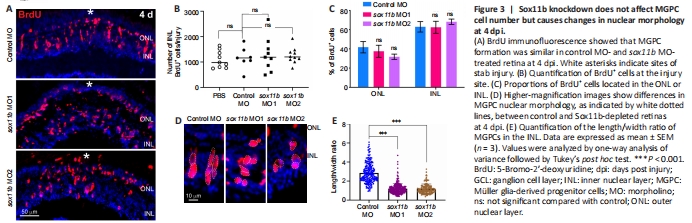
To investigate the role of Sox11b in MGPC formation, we electroporated a validated translation-blocking MO against Sox11b (sox11b MO1) (Veldman et al., 2007; Pillai-Kastoori et al., 2014) into the retina at the time of injury. Because most BrdU+ cells in the INL are MGPCs at 4 dpi (Fausett and Goldman, 2006), we quantified the number of these cells in the retina. BrdU immunofluorescence showed that Sox11b knockdown did not significantly affect the number of BrdU+ cells in the INL at the injury site compared to the control MO group (Figure 3A and B). The electroporation treatment did not significantly increase the numbers of INL proliferating cells, compared with the PBS control that lacked electroporation (Figure 3B), suggesting that electroporation itself did not cause extensive retinal damage. Notably, the electroporation of a second Sox11b translation-blocking MO (sox11b MO2) into the injured retina produced similar results (Figure 3A and B), confirming the above findings. Additionally, Sox11b knockdown did not alter the distribution of BrdU+ cells across the ONL and INL (Figure 3C). Although Sox11b knockdown did not affect MGPC formation, the nuclear morphology of MGPCs was altered in the INL (Figure 3A and D). Specifically, control MGPCs in the INL displayed an elongated, spindle-shaped nuclear morphology in which the long axis was largely perpendicular to the retinal layers (Figure 3D). In contrast, the nuclei of Sox11b-depleted MGPCs exhibited a more rounded shape (Figure 3D). Quantification of MGPC morphology revealed that Sox11b knockdown significantly decreased the length/width ratio for affected nuclei, compared with the ratio for control nuclei (Figure 3E). Because spindle-shaped MGPCs have been shown to migrate along the radial MG fibers to different retinal layers (Nagashima et al., 2013), the altered nuclear morphology observed in Sox11b-depleted MGPCs suggests that they exhibit defective radial migration.
Figure 4|Sox11b loss-of-function alters MGPC distribution across retinal layers after 7 dpi.
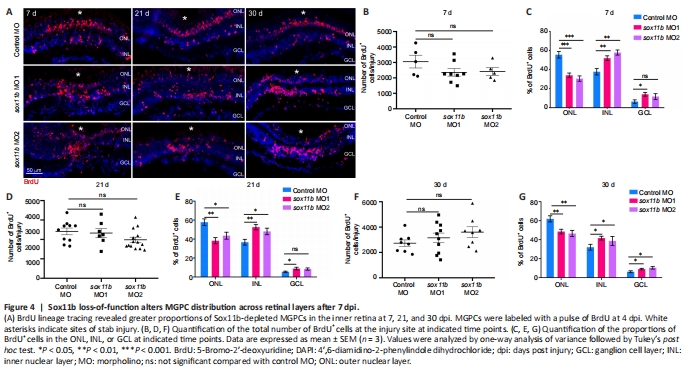
Because Sox11b exhibits late expression in MGPCs and has a nonessential role in MGPC formation, it may be involved in later stages of retinal regeneration (e.g., MGPC migration and fate determination). To explore this possibility, MGPCs were labeled with a pulse of BrdU at 4 dpi; the distributions of their progeny cells were examined at 7, 21, and 30 dpi. Consistent with the above findings at 4 dpi, Sox11b knockdown did not significantly affect the number of BrdU+ cells at 7 dpi (Figure 4A left panel, and 4B). However, assessment of BrdU+ cell locations revealed that Sox11b loss-of-function altered their distribution across retinal layers (chi-squared test, P < 0.05). In control retinas, BrdU-labeled nuclei were preferentially found in the ONL, rather than the INL and GCL, at 7 dpi (Figure 4A and C). In contrast, a greater proportion of the nuclei of Sox11b-depleted MGPCs was found in the INL, rather than the ONL (Figure 4A and C). Compared with control retinas, sox11b knockdown significantly reduced the proportion of MGPC nuclei in the ONL but increasing that proportion in the INL (Figure 4C). At 21 and 30 dpi, similar phenotypes were observed in retinas that had been treated with sox11b MOs (Figure 4A middle and right panels, and 4D–G). These results suggest that Sox11b regulates MGPC migration; Sox11b loss-of-function could lead to altered distributions of MGPCs at later time points.
Figure 5|Sox11b biases MGPCs toward a photoreceptor fate.
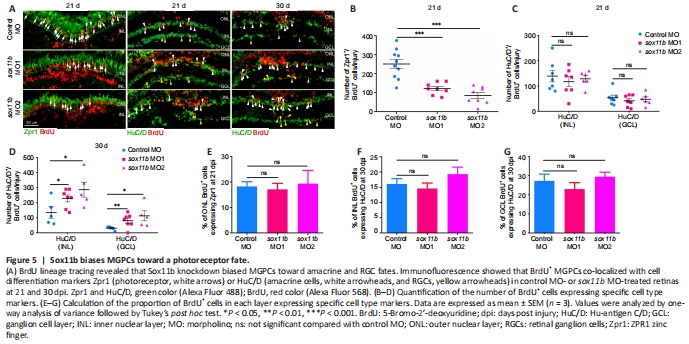
To investigate the role of Sox11b in MGPC fate determination, MGPCs were labeled with a pulse of BrdU at 4 dpi; their subsequent differentiation was examined by immunostaining with BrdU and cell type markers (Zpr1, photoreceptors; HuC/D in INL, amacrine cells; HuC/D in GCL, RGCs) at 21 and 30 dpi. At 21 dpi, Sox11b knockdown significantly reduced the number of BrdU+/Zpr1+ cells in the ONL (Figure 5A and B) but had no effect on the numbers of BrdU+/ HuC/D+ cells in the INL or GCL (Figure 5A and C); these results suggested that Sox11b promotes the differentiation of MGPCs into photoreceptors. Because Sox11b did not appear to affect amacrine cell or RGC regeneration at 21 dpi, while the proportion of MGPCs in the INL was greater in Sox11b-depleted retinas (Figure 4), we examined the differentiation of MGPCs at 30 dpi. Indeed, immunostaining revealed that Sox11b knockdown led to increased numbers of BrdU+/ HuC/D+ cells in both the INL and GCL (Figure 5A, and D). These findings suggest that the influence of Sox11b knockdown on amacrine cell or RGC regeneration occurred between 21 and 30 dpi, which is later than the influence of Sox11b knockdown on photoreceptors.
The similar effects of Sox11b silencing on MGPC distribution and neuronal regeneration across various retinal layers suggest that MGPC fates may be influenced by their location. Such influence would presumably cause the proportion of MGPCs that differentiate into each type of neuron within a single retinal layer to be comparable between controls and Sox11b morphants. Indeed, quantification revealed that the ratios of regenerated photoreceptors to ONL BrdU+ cells, amacrine cells to INL BrdU+ cells, and RGCs to GCL BrdU+ cells were all comparable between controls and Sox11b morphants (Figure 5E–G). Therefore, Sox11b may indirectly affect MGPC cell fate by regulating the radial migration and distribution of MGPCs.
Figure 6|Sox11b regulates the expression of Notch signaling components in injured retinas.
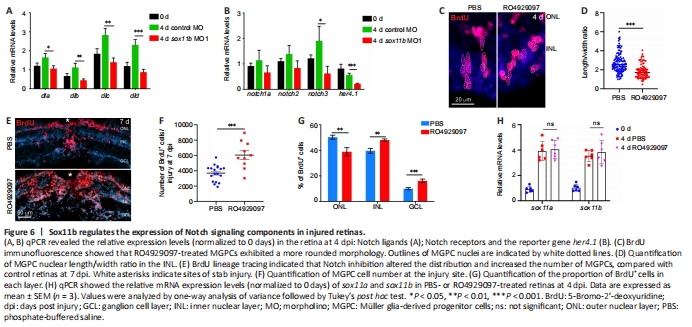
Notch signaling is an important regulator of neural progenitor proliferation and differentiation (Pinto-Teixeira and Desplan, 2014; Moore and Alexandre, 2020), as well as cortical neuron radial migration (Yang et al., 2017; Zhou et al., 2020b). Notch signaling components were reportedly induced in MGPCs, thereby regulating their differentiation, in injured zebrafish retinas (Wan et al., 2012). Because Sox11b loss-of-function resulted in defects in MGPC migration and fate determination, we examined its influence on the expression of Notch signaling components in the retina. At 4 dpi, qPCR revealed that Sox11b knockdown significantly reduced the expression levels of several Notch ligands, including dla, dlb, dlc, and dld; the Notch receptor notch3; and the Notch downstream target her4.1 (Wan et al., 2012) (Figure 6A and B). Thus, we inferred that Sox11b regulates Notch signaling in injured retinas. To investigate whether Notch inhibition might mimic the retinal phenotype of Sox11b morphants, zebrafish were intravitreally injected with PBS or the Notch inhibitor RO4929097 between 2 and 4 dpi; MGPC nuclear morphology was then examined via BrdU immunostaining. Indeed, Notch suppression altered MGPC nuclear morphology and decreased the nuclear length/width ratio in affected cells (Figure 6C and D), recapitulating the phenotype observed in Sox11b morphants. To examine the effect of Notch inhibition on subsequent MGPC distribution, MGPCs were labeled with a pulse of BrdU at 4 dpi; zebrafish were then treated with PBS or RO4929097 from 4 to 7 dpi. We found that Notch inhibition during this period led to an increased MGPC population (Figure 6E and F), consistent with the involvement of Notch in MGPC proliferation (Wan et al., 2012). Notably, Notch suppression resulted in greater proportions of MGPCs in the INL and GCL, compared with control retinas, while a smaller proportion of MGPCs was present in the ONL (Figure 6E and G); these results were similar to the findings in Sox11b-depleted retinas. qPCR analysis indicated that the retinal phenotype after Notch inhibition was not the product of altered Sox11 expression (Figure 6H). Taken together, these findings suggest that Sox11b regulates the expression patterns of Notch signaling components; it presumably controls MGPC migration and fate determination through this pathway.点击此处查看全文