视神经损伤
-
Figure 1|PABA has no effect on retinal cell death.
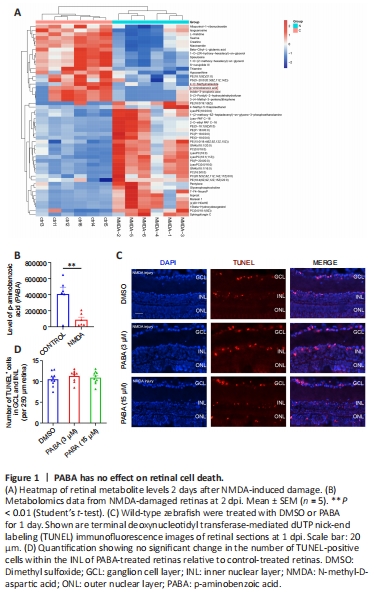
We performed untargeted metabolomic analyses to investigate alterations in metabolites in NMDA-damaged zebrafish retinas. Two days after NMDA-induced injury, we collected retinas and performed metabolome sequencing. The most significantly differentially expressed metabolites (increased and decreased) are listed in Table 2 and shown in the heatmap in Figure 1A. Compared with that in uninjured retinas, there was a significant decrease in PABA (P < 0.05; Figure 1B) in retinas following NMDA-induced injury.
To investigate whether PABA affects retinal cell death in zebrafish, we maintained zebrafish in water containing either DMSO or PABA and conducted TUNEL to quantify retinal cell death. At 1 day post-NMDA injury, no significant difference in the rate of cell death was detected between the PABA and control groups (P > 0.05; Figure 1C and D). NMDA-induced cell death in both groups was primarily localized to the ganglion cell layer (GCL) and inner nuclear layer (INL). These findings are consistent with previous studies showing that significant cell death occurred in the GCL and INL following NMDA injury, owing to the fact that NMDA receptors are mainly expressed by retinal ganglion cells and a subset of amacrine cells (Shen et al., 2006).
Figure 2|PABA promotes Müller glia dedifferentiation and division.
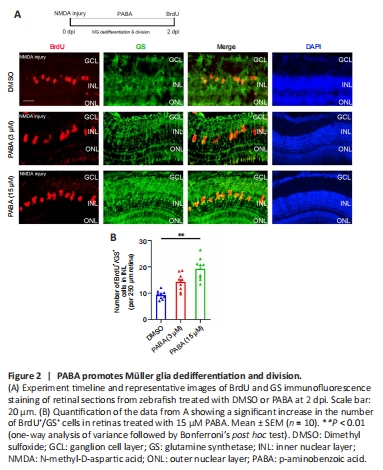
To investigate the effect of PABA on retinal regeneration, we first asked whether PABA contributes to Müller glia reprogramming. The initial steps involved in retinal regeneration in zebrafish include Müller glia dedifferentiation and asymmetric division to generate an MGPC (0–2 dpi), followed by rapid proliferation of MGPCs (2–7 dpi) (Fausett et al., 2008). We performed double immunofluorescence staining for BrdU and glutamine synthetase (GS) to identify proliferating Müller glia in the INL at 2 dpi and MGPCs in the INL at 4 dpi. In retinas treated with DMSO, few BrdU+/GS+ Müller glia were observed at 2 dpi (Figure 2A). In contrast, PABA (3 μM) treatment significantly increased the number of BrdU+/GS+ Müller glia, in a dose-dependent manner (Figure 2B), indicating that PABA promotes Müller glia proliferation. In subsequent experiments, we used 15 μM PABA. We did not observe retinal regeneration in an optic nerve crush model (Additional Figure 1), which is consistent with the results reported by Chen et al. (2022). They found no BrdU+ cells in the GCL of ONT+ retinae at 7?dpi, indicating that retinal ganglion cells had not yet regenerated.
Figure 4|PABA promotes MGPC proliferation.
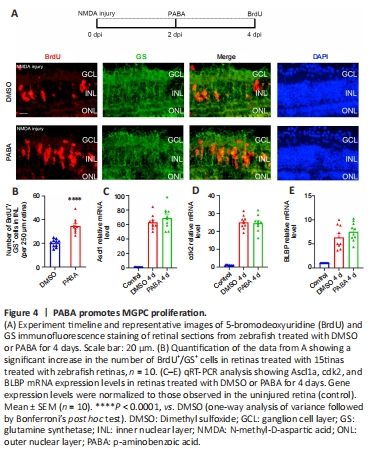
To study whether PABA plays a role in MGPC proliferation period, we examined BrdU+/GS+ cells in the INL at 4 dpi in zebrafish treated with PABA and DMSO. We maintained zebrafish in PABA and DMSO for 4 continuous days and detected MGPC proliferation in the INL at 4 dpi. The results showed that PABA treatment significantly increased the number of INL BrdU+/GS+ cells at 4 dpi compared with the DMSO group (Figure 4A and B), indicating that PABA promoted MGPC proliferation. qRT-PCR analysis showed that PABA treatment for 4 days had no effect on the expression of Ascl1a, cdk2, and BLBP compared with the control (Figure 4C–E).
Figure 5|Folic acid has no effect on Müller glia proliferation.
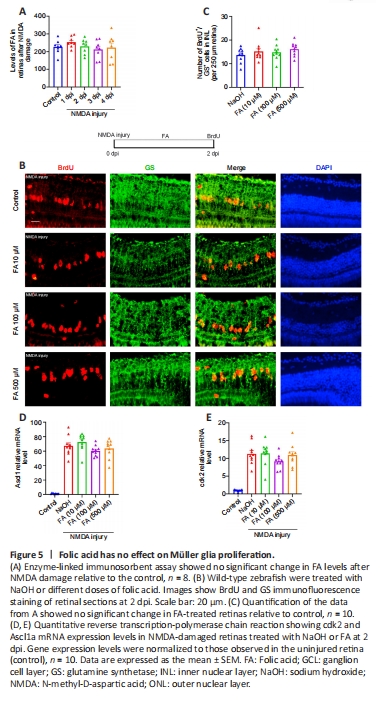
PABA is the precursor in FA biosynthesis, and FA is required to make DNA and RNA. We next asked whether PABA promotes retinal regeneration by increasing FA synthesis. First, we examined the FA concentration in retinas 1 and 4 days following NMDA injury. The results showed that there was no significant change in FA levels in the retinas after NMDA treatment (Figure 5A). To study the function of FA in retinal regeneration, we induced NMDA injury in zebrafish, treated the zebrafish with NaOH or FA, and performed BrdU immunofluorescence assay at 2 dpi. Our results showed that the FA group had a similar amount of BrdU+/GS+ cells as the control group, regardless of dose, at 2 dpi (Figure 5B and C), indicating that FA does not promote retinal regeneration. As for the mRNA expression levels of RAGs, FA administration had no effect on Ascl1a or cdk2 expression at 2 dpi (Figure 5D and E), in accordance with the BrdU immunofluorescence results. Taken together, these results indicate that FA does not promote regeneration of NMDA-damaged retinas and that PABA does not promote retinal regeneration by increasing FA synthesis.
Figure 6|PABA has no effect on microglial activation.
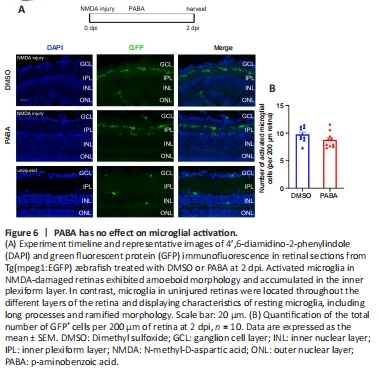
In the quiescent zebrafish retina, resting microglia are characterized by their long, irregular processes and fusiform morphology (Nagashima and Hitchcock, 2021). After NMDA-induced retinal injury in zebrafish, microglia migrate to sites of cell death, and their morphology changes rapidly and significantly from fusiform to amoeboid. Fluorescence microscopy showed that GFP+ cells accumulated in the inner plexiform layer (IPL) at 2 dpi (Figure 6A). The morphology of most of the GFP+ cells at the injury sites was amoeboid, suggesting that they were activated microglia. In contrast, the GFP+ cells in the uninjured group exhibited ramified morphology and were present in multiple different layers of the retina. There was no significant difference in the number of GFP+ cells between the PABA group and control, indicating that PABA did not promote microglial activation (Figure 6B).
Figure 7|PABA does not change MGPC distribution during differentiation.
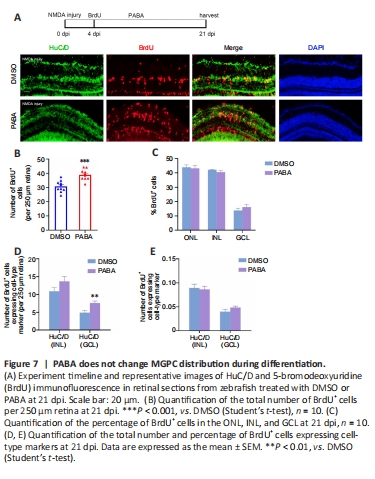
A previous study showed that MGPCs are intrinsically multipotent, migrate to any damaged nuclear layer, and differentiate into the appropriate cell type for that specific layer, although there is a proliferative bias in the most severely damaged nuclear layers (Powell et al., 2016). To assess the impact of continuous PABA treatment on MGPC distribution and retinal regeneration, we treated zebrafish with PABA for 21 days. Immunofluorescence showed that PABA treatment increased the number of BrdU+ cells at 21 dpi compared with the control (Figure 7A and B), suggesting that MGPC increased in response to PABA treatment. Compared with the control, PABA treatment did not significantly alter the percentage of BrdU+ cells in different retinal layers (Figure 7C), suggesting that PABA administration did not affect MGPC multipotency or distribution. In addition, staining for regenerated neurons, which were represented by BrdU+ cells expressing major cell-type markers INL HuC/D (amacrine cells) or GCL HuC/D (retinal ganglion cells) showed that there was no difference in the percentage of regenerated neurons between the PABA group and the control group (Figure 7E). In conclusion, PABA promoted MGPC proliferation but had no effect on MGPC distribution.
Figure 8|PABA promotes retinal regeneration in an Ascl1-dependent manner.
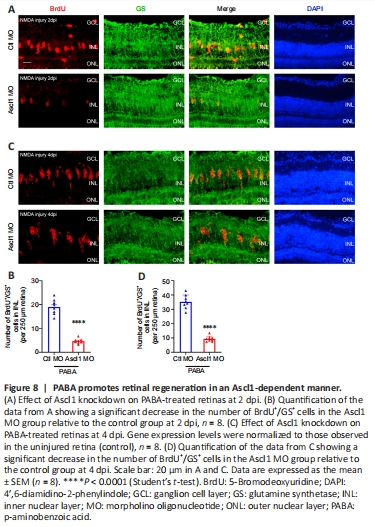
Our data suggest that PABA promotes retinal regeneration and increases Ascl1a expression. Because Ascl1a was previously shown to be a key factor in Müller glia dedifferentiation and MGPC proliferation (Ramachandran et al., 2010), we asked whether its induction in PABA-treated retinas was necessary for retinal regeneration. To test this, we performed gene knockdown of Ascl1a by intravitreally injecting a control or Ascl1a-targeting MO into PABA-treated zebrafish and labeled the proliferating Müller glia with BrdU at 2 or 4 dpi. The results showed that the Ascl1a-targeting MO significantly reduced the number of Müller glia by 80% in the PABA-treated retinas compared with the control MO at 2 dpi (Figure 8A and B). Furthermore, compared with the control MO, Ascl1a knockdown caused a significant reduction in MGPC proliferation, by 80%, at 4 dpi (Figure 8C and D). These findings are consistent with a previous study reporting that Ascl1a knockdown suppressed Müller glia division and MGPC proliferation in the injured retina (Ramachandran et al., 2010). These results indicate that PABA promotes retinal regeneration by increasing Ascl1a expression.