脑损伤
-
Figure 1|3′-Deoxyadenosine (3′-dA) reduces methamphetamine (METH)-mediated conditioned place preference (CPP) scores in mice and decreases hippocampal neuron activity.
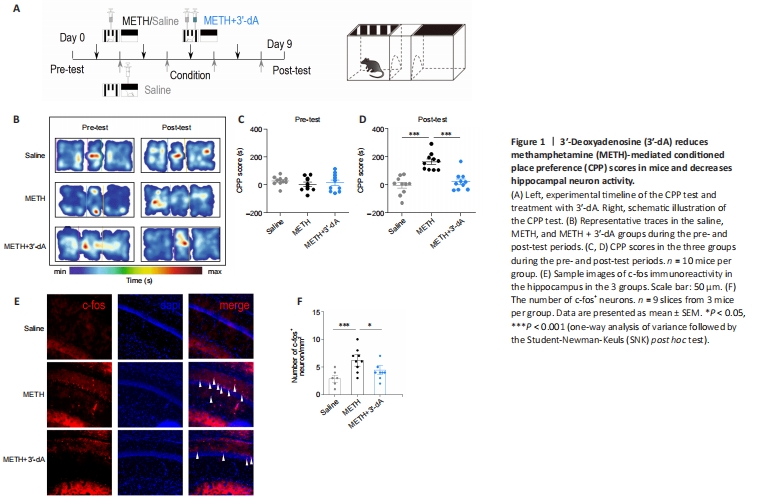
To determine whether 3′-dA plays a role in METH-acquired behavior, we treated 8-week-old C57BL/6 mice with 3′-dA prior to the CPP test (Figure 1A). The CPP scores of the METH group were significantly increased compared with those of the Saline group (P < 0.001). Meanwhile, the administration of 3′-dA decreased CPP scores in the post-test period (Figure 1B–D). Although the main region implicated in addictive memory is the nucleus accumbens, which receives dopamine inputs from the VTA via the mesolimbic pathway, we wanted to investigate associated changes in hippocampal neurons following METH-induced seeking behavior. Accordingly, we examined the expression of c-fos, a marker of neuronal activity, in the three groups (Chen et al., 2022). We found that the METH + 3′-dA group had fewer c-fos+ cells than the METH-treated group (P < 0.05; Figure 1E and F). These results demonstrate that 3′-dA improves METH-acquired behavior and decreases the activity of hippocampal neurons in mice.
Figure 4|3′-Deoxyadenosine (3′-dA) reduces methamphetamine (METH)-induced NOD-like receptor family pyrin domain containing-3 (NLRP3) activation and hippocampal neuronal damage.
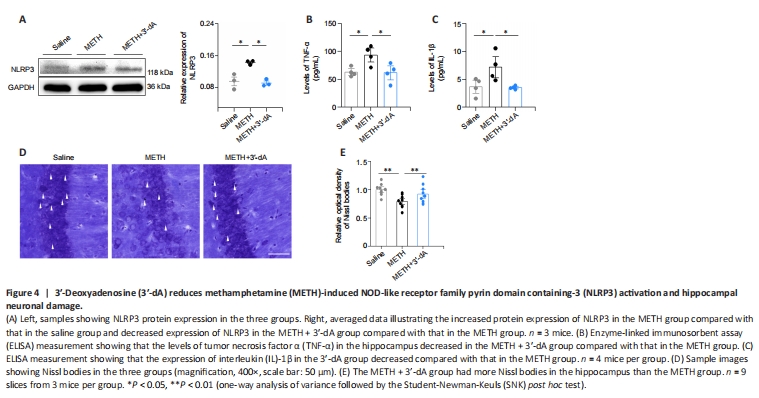
To investigate whether 3′-dA could attenuate neuroinflammation and neuronal damage in METH-addicted mice, we examined NLRP3 inflammasome expression and histological neuronal data in the three groups. We found that NLRP3 expression was higher in the METH group than in the Saline group, while treatment with 3′-dA reduced this elevation (P < 0.05; Figure 4A). Next, we extracted tissue supernatant to determine the relative expression of TNF-α and IL-1β. ELISA results showed that the relative expression levels of TNF-α and IL-1β were significantly higher in the METH group compared with the levels in the Saline group, while treatment with 3′-dA suppressed the elevation of these inflammatory factors (P < 0.05; Figure 4B and C). According to the Nissl staining data, 3′-dA attenuated the damage to neurons caused by METH administration (Figure 4D and E). Taken together, these results suggest that 3′-dA reduces METH-induced NLRP3 activation and IL-1β- and TNF-α-mediated inflammatory responses, thereby decreasing hippocampal neuronal damage.
Figure 5|NOD-like receptor family pyrin domain containing-3 (NLRP3) deficiency decreases methamphetamine (METH)-induced seeking behavior, reduces impaired synaptic plasticity, and prevents hippocampal neuronal damage.
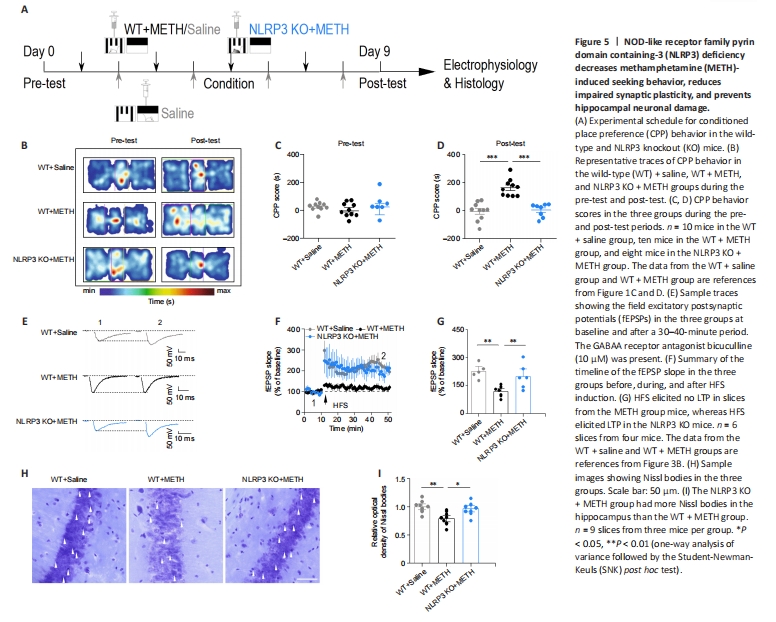
We have shown that 3′-dA decreased the expression of the NLRP3 inflammasome and aberrant synaptic plasticity in METH-exposed mice. Next, we asked whether the NLRP3 inflammasome contributes to aberrant synaptic plasticity, thereby affecting seeking behavior. We used NLRP3 knockout mice to verify the link between neuroinflammation and synaptic plasticity (Figure 5A). The three groups of mice were wild-type mice treated with saline (WT + Saline), wild-type mice treated with METH (WT + METH), and NLPR3 KO mice treated with METH (NLRP3 + METH). We found that CPP scores were higher in the WT + METH group compared with those in the WT + Saline group (P < 0.001). The CPP scores in the NLRP3 KO + METH group were lower than those in the WT + METH group, suggesting that a NLPR3 deficiency decreases METH-induced seeking behavior (P < 0.001; Figure 5B–D). Furthermore, we examined hippocampal synaptic plasticity in the NLRP3 KO + METH mice after METH administration. The results showed an increase fEPSP slope in the NLRP3 KO mice, suggesting that the impaired synaptic plasticity was restored in mice with a NLPR3 deficiency (P < 0.01; Figure 5E–G). Moreover, we collected brain tissue from the three groups of mice and conducted Nissl staining. We found that the damage to hippocampal CA1 neurons was attenuated in the NLRP3 KO mice compared with that in the WT+METH mice (P < 0.05; Figure 5H and I). Taken together, these data indicate that a NLPR3 deficiency reduces METH-related addictive behavior, neuronal damage, and altered synaptic plasticity.
Figure 6|An NOD-like receptor family pyrin domain containing-3 (NLRP3) agonist reverses the therapeutic effects of 3′-deoxyadenosine (3′-dA) in methamphetamine (METH)-treated mice.

To further determine whether 3′-dA exerts its therapeutic effect through the NLRP3 inflammasome, we applied nigericin (Nig), which is an NLRP3 agonist. We measured the CPP scores of mice given both Nig and 3′-dA (Figure 6A). There were no significant differences in CPP scores among the three groups during the pre-test (P > 0.05; Figure 6B and C). The CPP scores in the 3′-dA group were lower than those in the METH group (P < 0.001). The CPP scores in the METH + 3′-dA + Nig group were significantly increased compared with those in the METH+3′-dA group (P < 0.001; Figure 6D), suggesting that Nig reversed the effects of 3′-dA on METH-induced preference. The hippocampal field potential data showed that simultaneous administration of Nig, 3′-dA, and METH failed to induce LTP, suggesting that Nig reversed the recovery effect of 3′-dA on METH-induced synaptic plasticity (P < 0.01; Figure 6E–G). The Nissl staining data also demonstrated that Nig reversed the neuroprotective effect of 3′-dA in METH-addicted mice (P < 0.01; Figure 6H and I). Taken together, these data indicate that NLRP3 agonists can reverse the therapeutic effect of 3′-dA on METH administered mice, suggesting that the NLRP3 inflammasome mediates the role of 3′-dA in seeking behavior.