视神经损伤
-
Figure 5 | Inflammation regulates MG Jak1–Stat3 signaling in zebrafish retinas subjected to stab injury.
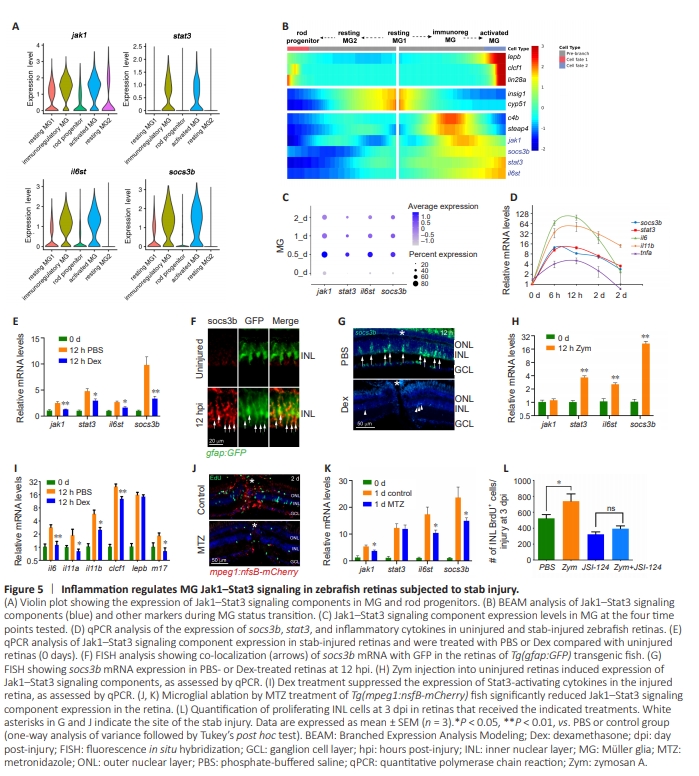
t has been demonstrated that Jak-Stat signaling is required for MG reprogramming and proliferation (Zhao et al., 2014). However, the connection between this signaling pathway in MG and inflammation remains unclear. We first examined the expression of Jak1–Stat3 signaling components in different MG subtypes and rod progenitor cells using our scRNAseq data. Our analysis showed that genes encoding key components of the Jak1–Stat3 signaling pathway, including jak1, stat3, il6st (GP130), and the downstream reporter socs3b, were enriched in immunoregulatory MG and activated MG, but not in resting MG1/MG2 or in rod progenitor cells(Figure 5A). Pseudotime analysis further demonstrated that expression of the genes encoding Jak1–Stat3 signaling components increased significantly during the transition from resting MG1 to activated MG, but decreased in the rod progenitor cell direction (Figure 5A and B). MG expression of genes encoding Jak1–Stat3 signaling components peaked at 12 hpi (Figure 5C, 0.5 days), coinciding with peak expression of typical inflammatory cytokines in the injured retina, as determined by qPCR (Figure 5D). Further examination of genes encoding Stat3-activating cytokines in the retina revealed that many of them, such as IL-4, IL-6, IL-11β, IL-21, and LIF (m17), were expressed by immune cells (Additional Figure 6A). Importantly, MG expressed genes encoding the corresponding receptors for some of these Stat3-activating cytokines (Additional Figure 6B and C). These findings suggest that secretation of Stat3-activating cytokines by immune cells could activate Jak1–Stat3 signaling in MG, thereby regulating MG reprogramming and proliferation. To investigate whether inflammation is required for activation of Jak1–Stat3 signaling in MG, zebrafish were treated with Dex or intravitreally injected with Zym to suppress or enhance retinal inflammation, respectively (Zhang et al., 2020; Zhou et al., 2022). qPCR analysis showed that Dex treatment significantly reduced the expression of genes encoding Jak1–Stat3 signaling component in the injured retina (Figure 5E). This was further confirmed by fluorescence in situ hybridization, which demonstrated that expressionof the Jak1–Stat3 reporter socs3b was increased in MG at 12 hpi (Figure 5F), while Dex treatment nearly abolished the socs3b signal in the inner nuclear layer (Figure 5G). Conversely, intravitreal injection of Zym into the uninjured retina led to a significant increase in retinal expression of Jak1–Stat3 signaling components (Figure 5H), suggesting that inflammation alone was sufficient to activate this signaling pathway in the intact retina. Furthermore, Dex treatment markedly reduced the injury-dependent expression of IL-6– family cytokines in the retina (Figure 5I), providing further support for the role of inflammation in regulating Jak1–Stat3 signaling in MG. Consistent with this, microglial ablation in Tg(mpeg1:nfsB-mcherry) transgenic fish (Zhang et al., 2020) significantly decreased the expression of Jak1–Stat3 signaling components in the injured retina (Figure 5J and K), indicating that microglia are necessary for activation of this signaling. We previously demonstrated that enhancing retinal inflammation via Zym injection promotes MG proliferation in the injured retina (Zhang et al., 2020; Zhou et al., 2022). Importantly, this effect was abolished in the retinas of fish treated with a Stat3 inhibitor JSI-124 (Figure 5L), suggesting that inflammation regulates MG proliferation through Jak1–Stat3 signaling.