神经损伤与修复
-
Figure 1|BrdU-immunolabeling in the cerebellar external granular of 10-day-old rats allowed to survive for 6 hours after a single injection of BrdU.
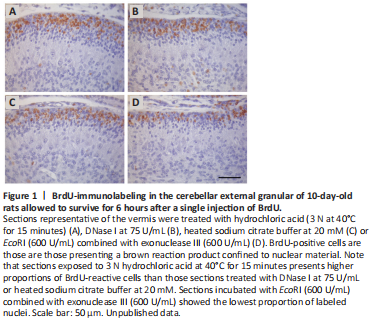
When these antigen retrieval procedures were systematically compared, it was found that the specificity of the staining pattern was affected by the antigen retrieval used (Dolbeare, 1995; Molina et al., 2017). In other words, these procedures give rise to considerable variability in the proportion of BrdU-stained cells. For example, protocols using 75 and 100 U/mL of DNase I, and 20 mM of heated sodium citrate buffer yielded similar numbers of BrdU-positive cells, although the values were smaller than those obtained with 3 N hydrochloric acid at 40°C during 15 minutes. 600 U/mL of endonuclease EcoRI combined with 600 U/mL of exonuclease III provided the smallest values. The reasons for these discrepancies are unknown, but they may derive from differences in the creation of single-stranded DNA. In this context, the disruption of the double-stranded DNA may be less accurate in those procedures using DNase I, heated citrate buffer or 600 U/mL of endonuclease EcoRI combined with 600 U/mL of exonuclease III, so that fewer antigenic sites can react with primary anti-BrdU antibody. Figure 1 depicts the variation in the number of labeled granule cell precursors in the cerebellar external granular layer among sections treated with several antigen retrieval solutions, such as hydrochloric acid (3 N at 40°C for 15 minutes) (Figure 1A), DNase I at 75 U/mL (Figure 1B), heated sodium citrate buffer at 20 mM (Figure 1C), and EcoRI (600 U/mL) combined with exonuclease III (600 U/mL) (Figure 1D). Taken together, these results have suggested that, depending on the antigen retrieval solution, the detection threshold for the BrdU incorporated into replicating DNA may be different.
Figure 2|Light photomicrographs illustrating ectopic granule cells from rats administered with bromodeoxyuridine on embryonic days 13–14 and sacrificed on postnatal day 90.
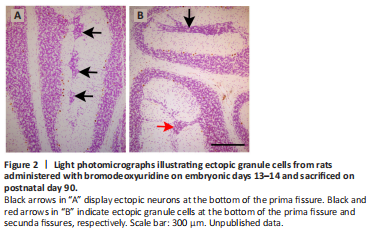
When the toxicity of repeated BrdU administrations on the proliferating neuroblasts are analyzed, the data are controversial. This is because previous reports have indicated that the administration of 50 mg/kg of BrdU in a cumulative labeling sequence during the prenatal life, is well tolerated and produces neither cytotoxic effects nor alters, at embryonic day 14, cell cycle dynamics in the dorsomedial region of the mouse cerebral wall (Takahashi et al., 1992, 1993, 1995a). No alterations have been reported when a succession of BrdU injections were administered on each of embryonic days 11 to 17 (Takahashi et al., 1995b). In line with these data, it has been shown that, in a cumulative BrdU (35 mg/kg) labeling sequence, no cytotoxic effects were observed in the rat cerebellar neuroepithelium (Martí and Rodríguez-Vázquez, 2020). In contrast, when several injections of BrdU were administered at 8-hour intervals over 2 successive days between days 11 and 21 of rat pregnancy and the offspring was analyzed in adulthood, defects in the proliferation, migration, and settling of the Purkinje cells were seen. Moreover, reductions in the sizes of the cerebellar cortex and deep nuclei, as well as foliation defects were found. Interestingly, the effects BrdU varied depending on the age of the animals (Sekerková et al., 2004b). In this line, results from my laboratory have revealed that when rats were administered with BrdU on embryonic days 13 and 14, following the abovementioned progressively delayed cumulative method, several granule cells were arrested in the molecular layer where they formed an ectopic zone. Examples of clusters of ectopic granule cells located in the bottom of the prima and secunda fissures are shown in Figure 2.