NRR:中国华中科技大学附属同济医院王伟教授团队认为脊髓V3中间神经元在运动控制中具有重要的作用
撰文:章若樱
脊髓V3中间神经元属于兴奋性谷氨酸能神经元,广泛分布于脊髓的背侧、中间及腹侧区域。它们在中枢神经系统中参与构建广泛的神经回路连接,并在运动功能中发挥关键作用,特别是在行走过程中维持稳定且协调的步态。更为重要的是,在脊髓损伤发生后,这些神经元能够保持其电生理兴奋性,并促进本体感觉信号向运动神经元的有效传递,这对于启动类似行走的复杂、协调性以及节律性活动具有重要意义。因此,脊髓V3中间神经元被认为是促进脊髓损伤后运动功能恢复的潜在治疗靶点。对于解析各类脊髓中间神经元以及靶向不同中间神经元治疗脊髓疾病是目前研究的热点之一,特别是在近2年有了新的进展。但是,目前针对脊髓V3中间神经元治疗脊髓疾病的治疗策略还非常有限。
中国华中科技大学附属同济医院王伟教授团队在《中国神经再生研究(英文)》(Neural Regeneration Research)上发表了题为“Roles of spinal V3 interneurons: Roles in controlling movement in
healthy and injured conditions”的综述,文章首先总结了脊髓V3中间神经元不同亚型的解剖分布和生理学特征(图1),其次总结了脊髓V3中间神经元在生理状态下对不同运动模式的调控和在脊髓损伤后神经元环路的可塑性(图2),并且将不同V3亚型和不同的轴突投射模式以及对应的功能联系起来(图3)。最后重点讨论了脊髓V3神经元在脊髓损伤疾病中可能存在的治疗靶点(图4)。了解脊髓V3神经元在生理和病理下的作用有助于制定治疗脊髓疾病的潜在治疗策略。
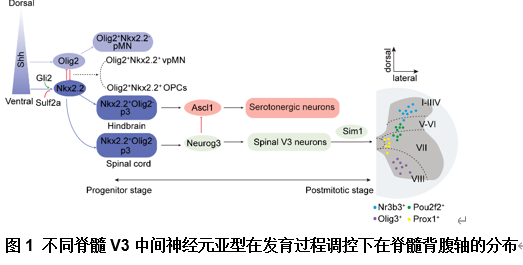
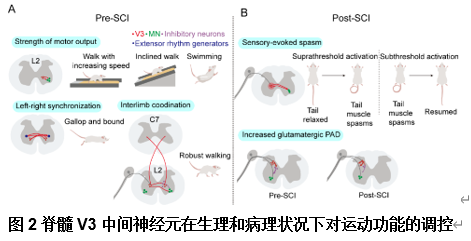

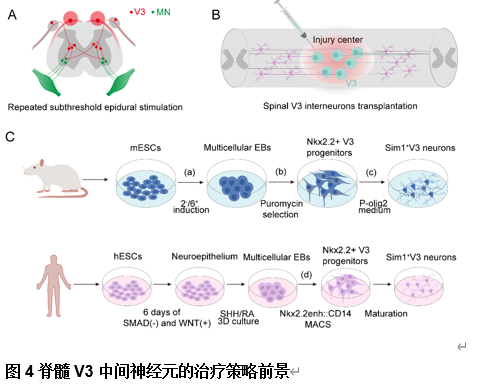
王伟等认为,脊髓V3中间神经元是一个高度异质性的神经元群体,广泛分布于成熟脊髓的背腹轴和头尾轴。尽管脊髓V3中间神经元在整体上对维持稳定且协调的运动功能具有重要作用,然而其不同亚型在运动调控中表现出各自特异的功能特征[1]。这一现象与成年斑马鱼中不同脊髓V2a神经元亚型的研究结果相一致,后者提出不同V2a神经元参与到不同的神经环路中,参与调节不同的运动频率 [2]。然而,目前对于脊髓中不同V3神经元亚型的具体功能尚未完全阐明。近年来,研究已构建了多种小鼠和人类脊髓神经元亚型基因表达的图谱,涵盖发育期 [3, 4]及成年期[5, 6]的小鼠模型,以及发育期[7, 8]和成年期 [9, 10]的人类样本。借助单链RNA测序与空间转录组学技术,已识别出多种具有不同空间分布特征的神经元簇。因此,未来研究应重点聚焦于在人脊髓中V3中间神经元及其亚型的鉴定,以及深入理解其在神经环路构建及运动行为调控中的具体功能,这对于推动脊髓疾病的机制研究与治疗策略制定具有重要意义。此次综述还提出了未来的研究方向和研究重点, 包括(1)对于脊髓不同节段分布的脊髓V3中间神经元功能的探索,如分布于脊髓颈段的V3神经元是否参与到前肢运动功能的调控中。(2)未来研究应拓展脊髓V3中间神经元参与到感觉神经环路中的研究,因为脊髓损伤后除运动功能的恶化,还伴随着异常疼痛,痉挛和瘙痒的发生,而脊髓V3神经元作为兴奋性中间神经元是否参与到感觉环路的调控也是研究的重点方向,未来的研究应拓展中间神经元在脊髓感觉-运动环路中发挥的重要作用。(3)目前脊髓疾病的研究聚焦于实现脊髓损伤后神经环路的激活和重建,例如对脑和脊髓的电刺激以及干细胞移植等,这些治疗方法固然有效,但其针对的脊髓神经环路并不具备特异性,因此未来的研究重点仍在于采用单细胞测序、空间转录组学等方法对单一脊髓神经元进行详细的解析,包括其特异性标志物以及控制其再生的信号通路。这为未来靶向不同脊髓中间神经元生长至特异性靶点,构建控制不同功能的特异神经环路发挥治疗作用提供了有利的证据,而对于脊髓神经元的解析也将推动其他包括药理学,光遗传学等更多治疗策略的发展。
综上所述,目前已有越来越多的研究致力于识别脊髓V3中间神经元以及其在运动、感觉和自主神经系统功能中的作用。随着基因编辑与病毒示踪技术的发展,科研人员对该类中间神经元群体的认识日益深入。然而,关于脊髓V3中间神经元的分子特征、功能特性及其在脊髓损伤后的潜在治疗效应,仍存在诸多尚未明确的问题。因此,未来研究亟需进一步对其本身及其亚型进行系统性表征,并加深对其在脊髓损伤后神经功能重塑中所扮演角色的理解。唯有如此,才能为开发具有靶向性的治疗策略奠定基础,从而有效挖掘其在神经修复中的治疗潜力。
#br#
原文链接:https://doi.org/10.4103/NRR.NRR-D-25-00191
参考文献#br#
[1] Borowska J, Jones CT, Zhang H, et al. Functional subpopulations of V3 interneurons in the mature mouse spinal cord. J Neurosci. 2013;33(47):18553-18565.#br#
[2] Ampatzis K, Song J, Ausborn J, et al. Separate microcircuit modules of distinct v2a interneurons and motoneurons control the speed of locomotion. Neuron. 2014;83(4):934-943.#br#
[3] Delile J, Rayon T, Melchionda M, et al. Single cell transcriptomics reveals spatial and temporal dynamics of gene expression in the developing mouse spinal cord. Development. 2019;146(12):dev173807.#br#
[4] Osseward PJ, 2nd, Amin ND, Moore JD, et al. Conserved genetic signatures parcellate cardinal spinal neuron classes into local and projection subsets. Science. 2021;372(6540):385-393.#br#
[5] Russ DE, Cross RBP, Li L, et al. A harmonized atlas of mouse spinal cord cell types and their spatial organization. Nat Commun. 2021;12(1):5722.#br#
[6] Sathyamurthy A, Johnson KR, Matson KJE, et al. Massively parallel single nucleus transcriptional profiling defines spinal cord neurons and their activity during behavior. Cell Rep. 2018;22(8):2216-2225.#br#
[7] Shi Y, Huang L, Dong H, et al. Decoding the spatiotemporal regulation of transcription factors during human spinal cord development. Cell Res. 2024;34(3):193-213.#br#
[8] Li X, Andrusivova Z, Czarnewski P, et al. Profiling spatiotemporal gene expression of the developing human spinal cord and implications for ependymoma origin. Nat Neurosci. 2023;26(5):891-901.#br#
[9] Yadav A, Matson KJE, Li L, et al. A cellular taxonomy of the adult human spinal cord. Neuron. 2023;111(3):328-344.e327.#br#
[10] Zhang D, Chen Y, Wei Y, et al. Spatial transcriptomics and single-nucleus RNA sequencing reveal a transcriptomic atlas of adult human spinal cord. Elife. 2024;12:RP92046.#br#
#br#